|
Authors: Claude Bajada, Nils Muhlert, Ilona Lipp Infographic: Roselyne Chauvin Expert editors: Alfred Anwander, Jurgen Gatt Newbie editor: Caroline Jantzen Introduction Neuroanatomy is one of the most exciting topics in neuroscience! Some readers may disagree, but for now, humor us and read along. With the help of this On-Demand post, we will convince you not only that anatomy is a useful endeavour but that it is one where much beauty is found. Our journey starts with the fundamental notion that the structure and the function of objects are tightly coupled; sometimes in ways that are not obvious. Understanding the complexity of the brain’s structure, hopefully, allows researchers to build more accurate models of brain function. Neuroscience has however become such a transdisciplinary subject that it is not unexpected to meet a top scientist who has never seen a cadaveric brain. Indeed, while most neuroscientists have acquired a basic understanding of brain anatomy, learning about the main gyri, sulci and nuclei, few remember the function or location of the mysterious substantia innominata or of the periaqueductal grey. If you are one of such scientists, fear not, you are not alone. Neuroanatomy is a vast and somewhat arcane subject, steeped in history. As such, for the sake of our sanity and yours, in this singular blogpost we decided to restrict our dealings to the topics that are most commonly tackled within the neuroimaging community. This is an overview of the major landmarks and structures that one can expect to see on an MRI scan and the ongoing conundrum of how to subdivide the brain into useful (sub)regions for further analysis - parcellation (See OHBM How-To Machine Learning on performing data-driven parcellation of MRI data). Why is understanding anatomy important? Getting from MRI DICOM files to a statistical map of significant clusters that shows differences between two experimental groups requires no anatomical knowledge whatsoever. But coming up with sensible brain based hypotheses, interpreting findings and communicating them to your fellow colleagues relies on a common language in the field: anatomy. In his video, David Van Essen (from min. 0:50) details the basic features of human brains, such that we have two hemispheres of about 1000 cm2 volume and a 2-4mm thick cortex that is highly convoluted. While providing an overview of the developmental mechanisms leading to the adult brain as we know it, he points out the huge individual variability in brain anatomy that requires us to apply flexible approaches to neuroimaging studies. These approaches include advanced image registration and the use of atlases, but also functional localisers, individual tractography, individual parcellations and last, but not least, knowing our anatomy. A knowledge of anatomy can make your life as a neuroscientist a lot easier. For example, it can aid placement of volumes of interest in MR spectroscopy, it can allow you to evaluate the output of automated segmentation pipelines, and, of course, help you quickly identify patterns of activity before receiving confirmation from atlases. Interested in doing some fancy high field, layer-specific, fMRI (see Noam Harel’s video)? Anatomical knowledge is vital! Also, recent evidence suggests that it can help increase the reproducibility of findings between labs, by improving accuracy and reducing noise when carrying out tractography in diffusion MRI. Understanding anatomy has clear implications for neuroimagers! Nomenclature, Etymology and Orientation in the Brain One of the first battlegrounds for the new student of anatomy is understanding the cryptic vocabulary of experts. Throughout the videos presented in the OHBM on-demand anatomy courses, various experts refer to different parts of the brain on the assumption that we all share a similar understanding of the language. To bring everyone to the same page, we will review the major terms that are crucial to understand before diving into any neuroimaging study. Anatomy uses many words borrowed, butchered, and stolen from Latin and Greek. Cerebrum is Latin for “that which is carried toward the head.” Cephalon is ancient Greek for head. Encephalon is the substance that is found inside the head, the cauliflower-resembling organ we now know as the brain. Any word encountered that has elements of these words refers to the brain. For example, the word cerebellum is the diminutive of cerebrum. The diencephalon (across the brain) is made up of the thalami (chambers), hypothalamus (below the thalamus) and epithalamus (above the thalamus). Knowing the etymology of the words makes remembering these ludicrously named structures easier. Readers are referred to this amusing article for more. Orientation is also often done in Latin. In their talks, Svenja and Julian Caspers regularly use two different ways of describing orientation in the brain. The terms superior, anterior, inferior and posterior (SAIP) versus the terms dorsal, ventral, rostral, and caudal (DVRC). While elite neuroanatomists would have no difficulty using these terms, for the neophyte they can be confusing. They actually refer to two completely independent coordinate systems. The SAIP approach is a real world orientation system. The terms themselves are intuitive and most people need little more explanation than the terms themselves. DVRC is another story! This is an orientation system that depends on the organism itself, the terms relate to parts of the body, once again, in Latin. Dorsal means towards the back, ventral is towards the belly, rostral is towards the beak (or nose), while caudal is toward the tail. Our bipedal nature makes this orientation system unintuitive - at the level of the spine, brainstem and cerebellum, rostral means towards the top of our body (upwards towards the nose), but in the cerebrum, rostral means towards the front of our body (forwards towards the nose). If it is still murky in your heads we would advise dipping into an introductory neuroanatomy textbook for some pretty pictures of the two orientation systems. When you hear the term medial, this means “towards the middle of the brain”, whilst lateral indicates “towards the sides”. 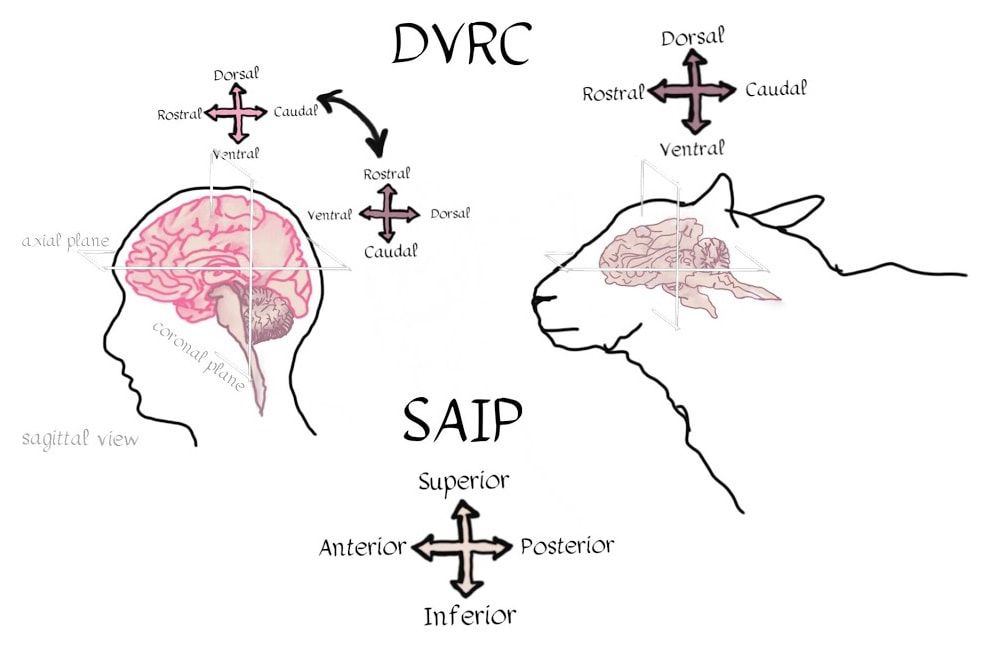 The frames of reference in neuroanatomy change for the cerebrum compared to the cerebellum, brainstem and spinal cord. For quadrupeds, like sheep, no such change is seen. Walking on two feet is great for reaching things, but not so great for keeping neuroanatomy simple! Another important set of terms relate to the way one can slice a brain. This is generally what we do when viewing MR images. Axial, or horizontal, slices allow you to scroll through the brain from top to bottom through the axis of the brain; sagittal slices from left to right derived from the latin word for arrow (think of the way an archer holds their bow and arrow). Finally, coronal sections provide a view of the brain as if it were cut through with a burning hairband shaped crown from ear to ear, slices moving anterior to posterior. (Yawn) Thank you for that primer, but when I look at an MRI scan, I still feel completely lost!! How do I get my head around it? MRI scans are tricky, they are often viewed as two dimensional slices and depending on the cut, and on the individual, it can still be very difficult to orient yourself, especially when the person was not lying straight in the scanner. As Svenja states in her video (min. 1:20), there are some general organising gross anatomical features that remain relatively consistent across individuals. These are called landmarks. So, what are the major landmarks in the brain? Some aspects of anatomy are pretty clear. Every healthy human brain has a cerebellum, brainstem and cerebrum... and a substantia innominata, of course. It has ventricles filled with corticospinal fluid, white matter that looks homogeneous on a conventional MRI scan, easily spottable subcortical gray matter nuclei, and cortical gray matter. Looking at a whole human brain from the outside shows a cortical folding pattern with specific sulcal and gyral structures, which have been labelled and can be used to orient yourself around the occipital, temporal, parietal and frontal lobes. In his video, Julian explains the main sulcal and gyral landmarks and how to find them in structural MR images. For example, if you spot an ‘M’ and ‘U’ in a lateral sagittal section, you have most likely found the central sulcus and precentral sulcus (from min. 11:36). An omega sign in a superior axial section indicates that you have found the motor cortex, while the cingulate sulcus appears as a bracket sign (from min. 14:40). The Figure below summarizes Julian’s guide to landmark spotting strategies. 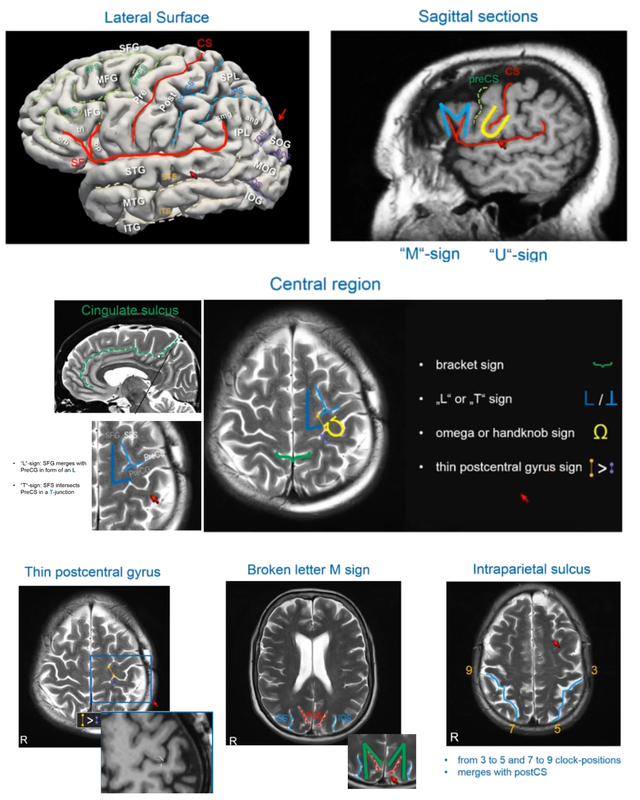 These landmarks are definitely useful, but they seem quite vague for reporting the spatial locations of my findings! A wise anatomy professor once complemented one of the authors of this piece (CJB) by stating that he was the owner of a “gelatinous mass of a brain.” Despite the gyral formations, the brain does indeed look like one amorphous clump of jelly. Notwithstanding the repeated news headlines claiming that Neuroscientists have found the region of the brain responsible for X, it is notoriously difficult to consistently define brain regions across different individuals. While the macrostructure of the brain, such as the main sulcal and gyral pattern, is useful to orient yourself on a whole brain or MRI scan, its macrostructure does not necessarily relate very well to the underlying brain function, which might be more closely related to the neuronal microstructure of a cortical brain area. To decide how to define brain areas based on cell anatomy, we first need to think about what neural features to use in order to separate them (also see this paper for cortical parcellations). Academic Journal Articles often refer to specific brain regions such as Brodmann area 17. Is this parcellation based on neural features? Yes! During the late 19th and early 20th century, anatomists started discovering that, while the cortex looks fairly homogeneous to the naked eye, it consists of various layers that differ in their cell type, cell composition, and function. Anatomists such as Cecile and Oskar Vogt, Constantin von Economo, and Korbinian Brodmann spent their time observing microscopic features of the cerebral cortex and classifying it according to similarity. These areas have become known as parcels. Undoubtedly the most famous parcellation scheme is Korbinian Brodmann’s 1909 atlas. In her video, Nicola Palomero-Gallagher shows some of the main historical cortical parcellations. She points out that the parcellation you get depends largely on how boundaries are defined. She then explains how more quantitative and objective approaches are used for not only finding parcellations in individual post-mortem brains, but also how this can be taken further into generating population maps that reflect individual variability in the boundaries of the areas. Cytoarchitecture is not the only feature that was used to parcellate the brain. At the same time that Brodmann was using cytoarchitecture, the Vogts tended to use myeloarchitecture to define regions. In his video, Matt Glasser explains how myelin-sensitive MRI contrasts can be used to study cortical myeloarchitecture and how that helps align cortical surfaces across individuals. But there is no reason to simply stop there, why not use the distribution of receptors for neurotransmitters to delineate brain areas?  In fact, while the cyto- and myeloarchitecture of the cortex tells us something about the type of processing happening in a cortical region, neuroreceptor density can also tell us quite a bit about different neural functions and how they become impaired in disease states. However, as Karl Zilles explains in his video, while cortical regions differ in their receptor fingerprints, there does not seem to be a clear relationship between the parcellations based on cyto- and myeloarchitectonics and those based on neurotransmitter receptor maps. 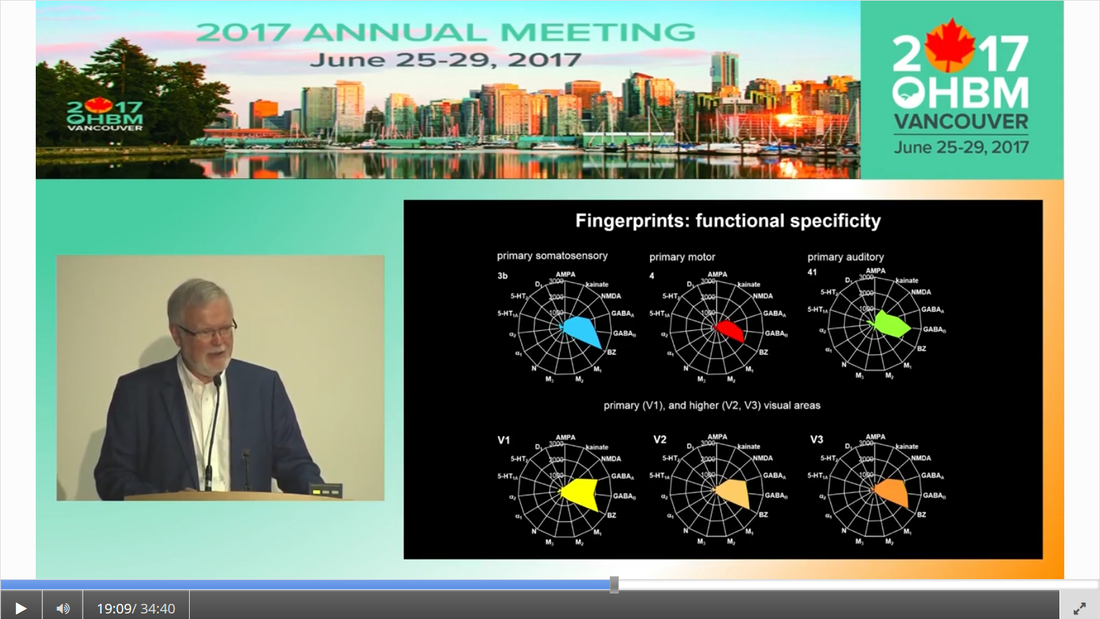 How can I link my imaging research to the histological parcellation of the brain? Functionally, the cortex is often divided based on the order of information processing: primary sensory areas are the first ones to receive sensory information, secondary areas do further processing, and association areas integrate information from different sensory modalities. Some functional units clearly match the microstructural organisation of the brain. For this reason, Brodmann’s atlas is often used to report the location of activation in functional imaging studies. Particularly high correspondence between function and microstructure has been reported for primary areas. In her video, Katrin Amunts explains how to identify the primary areas cytoarchitectonically, including the primary motor cortex (from min. 3:00), the primary auditory cortex (from min. 8:20), the primary visual cortex (from min. 9:20), and Broca’s area (from min. 12:37). However, how well can these cytoarchitectonically distinct areas be localised based on the anatomical landmarks visible on conventional MRI scans differs. In her video, Nicola Palomero-Gallagher provides examples for brain regions where the gyrification patterns nicely coincide with the microstructure and where it does not. Practical examples of how to make use of the correspondence in the visual system are given by Kalanit Grill-Spector in her video. She explains the anatomical localisation and microstructural features of place-selective regions within the so-called collateral sulcus (from min. 5:30) and face-selective regions in the so-called mid-fusiform sulcus (from min. 09:53). (For a good overview of the visual system also see Rainer Goebel’s video (from min. 3:12)). Primary areas are also characterized by specific mesoscopic organisation called topography (explained in Daniel Margulies’s video from min. 1:10). How high resolution fMRI can be used to study such organisation is explained by Rainer with examples of retinotopic mapping (min. 6:30). In order to be able to spatially localize results from your imaging study to parcellations based on the underlying cortical microstructural profile, findings from detailed postmortem characterisation have to be somehow transformed into usable atlases for in vivo imaging. In his video, Simon Eickhoff explains how probabilistic cytoarchitectonic mapping based on large-scale histology can aid with the spatial identification of MRI findings (from min. 7:28). He also goes into details on how to practically go about the question “Where is my blob?” (from min. 14:00). But how many brain regions are there now and how should I define them? How many regions there are depends on how you parcellate the brain (indeed experts often can’t agree on how many lobes there are! To limbic, or not to limbic?). To aid the localisation of findings and the definitions of regions of interest, brain atlases have been created. These atlases represent parcellations of a representative template brain, made to help you define your regions. When using these atlases (described in more detail here), we need to understand where they come from and what their limitations are, to decide which is the best atlas for our purpose. The different available atlases are based on various features of the brain (e.g. see Paula Croxson’s video on parcellation based on histological and microstructural features or Danilo Bzdok’s video on functional parcellations), and as you will find out there is no simple way of defining ‘brain regions’. There is also no reason to restrict oneself to a single feature of interest. Multimodal parcellations are becoming more popular! In her video, Paula Croxson explains that a robust parcellation of the brain has various advantages, such as help with localisation of function and also for understanding individual variation. In some contexts, parcellations into individual brain regions may not even be the way to go. For example, higher cognitive functions rely on large-scale networks and a complicated interplay of different regions. Functional connectivity is often done to tap into these networks (also see On Demand post about that). In his talk, Daniel explains how some local changes in functional connectivity even correspond to cytoarchitectonic boundaries (from min. 7:14). Finally, of course, there is no cortical hegemony in the brain, even though reading the neuroimaging literature seems to imply it. Hence, all the concepts and approaches discussed for cortical parcellations also apply for subcortical parcellation. We have discussed the cortex and the grey matter, can you tell me something interesting about white matter anatomy? Of course! Like everything else in anatomy, to speak about white matter deserves a little bit of time travel to the nineteenth century. This is the era where all (or most) of the “great tracts” were first described. We say described rather than discovered because there is nothing intrinsic to a tract that requires it to be so! This century (and parts of the previous) is the home to German and French giants such as Johann Christian Reil (did you know that the insula is sometimes referred to as the “Island of Reil?”, which is the word for island in Latin), who first described the arcuate and uncinate fasciculus, Karl Friedrich Burdach who identified the inferior longitudinal fasciculus, as well as Joseph and Augusta Dejerine, and Heinrich Sachs who all made contributions to confusing and contorted the white matter lexicon that we all currently know and love. A good resource on white matter tracts is this paper. In his video, Marco Catani gives an excellent introduction to the different types of white matter tracts that you may encounter during your research (from min. 1:30). He explains that ascending and descending projection fibres connect subcortical with cortical regions, that commissural fibres connect left and right hemisphere, and that association fibres serve feedforward and feedback connections. He also goes into detail about how to evaluate the anatomical plausibility of diffusion MRI tractography, which is currently the only approach that we have to investigate white matter non-invasively (See OHBM How-To Diffusion MRI). If you are specifically interested in the white matter tracts of the occipital lobe, Svenja’s talk guides you through this area of the brain. She goes into details on projection fibres (from min. 1:30), such as the optic radiation, association fibres (from min. 9:27), such as the inferior longitudinal fasciculus, the inferior fronto-occipital fasciculus and the superior longitudinal fasciculus, intralobar association fibres (from min. 16:15) and transcallosal fibres (from min. 21:00). Can we assign functions to white matter? No and yes. Tracts are not really thought of primarily as processing regions, therefore the naive labeling of tracts with specific functions may be misguided. However, knowing which tracts tend to be associated with certain functions (particularly their disruption secondary to damage - a disconnection) is crucial for any neurosurgeon’s work. In his video, Hugues Duffau explains this beautifully. He describes the fascinating procedure behind intraoperative direct electrical stimulation of white matter (from min. 3:00) and the way that this is used in neurosurgical mapping. Also check out this paper on disconnections and dysfunctions. Where is anatomy research heading with MRI developments? While anatomy seems like an old thing, there is still a lot we haven’t agreed on. With developments in high resolution imaging (described in Noam’s video), we have new ways of understanding how the brain is structured. Noam gives an overview of how moving to high field strength allows to obtain images with higher resolution and more sensitive studying of functional anatomy. Focussing on the visual system, Rainer gives examples for specific developments in ultra-high-resolution functional imaging that allow to study the meso-scopic functional organisation of neurons in vivo. 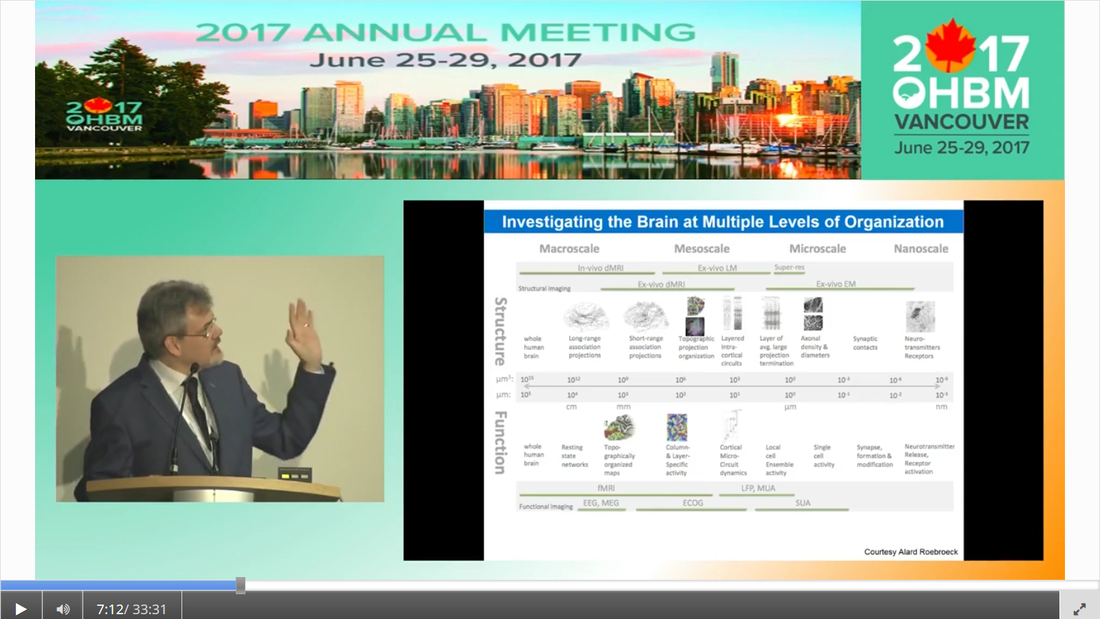 While many functional imaging findings have been superseded as the resolution and complexity of processing improve over the decades, neuroanatomical findings tend to be less dependent on the specific technique. As Marcel Mesulam said in an interview with the OHBM Blog: “The beauty about neuroanatomy is that it changes over millions of years. So once you discover something, it’s true for a few million years. And I have made some discoveries in neuroanatomy that were published maybe 30 to 35 years ago and are as true today as they were then.”
0 Comments
Your comment will be posted after it is approved.
Leave a Reply. |
BLOG HOME
Archives
January 2024
|
 RSS Feed
RSS Feed